What is Regeneration?
Regeneration is one of the processes in which if an organism is cut into several pieces, each of its parts regrows to the original state. This process is carried out by specialized cells called stem cells. It takes place in organisms that have a very simple structure with very few specialized cells.
The cells divide quickly into a large number of cells. Each cell undergoes changes to form various cell types and tissues. This sequential process of changes is known as development. The tissues form various body parts and organs.

Types of Regeneration
There are two types of regeneration:
i) Morphallaxis
This type of regeneration has little growth and depends upon tissues repatterning. For eg., Hydra grows by loss of cells from its end and by budding. When cut into two, the upper part develops into foot while the lower part develops into the head. The formation of nearby heads is inhibited by the head region.
ii) Epimorphosis
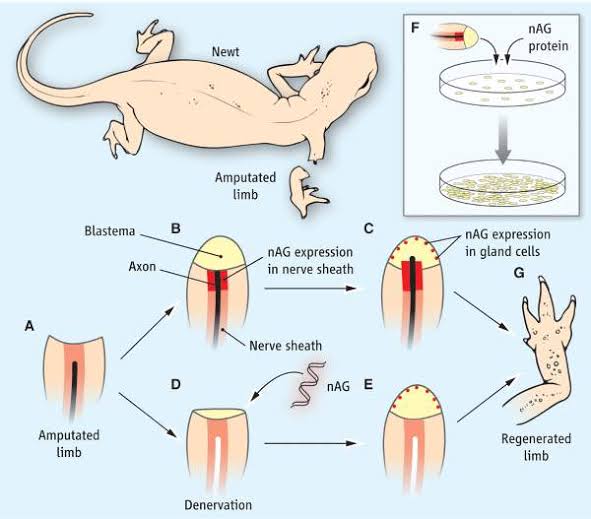
This type of regeneration depends upon the growth of new and properly patterned structures. For eg., regeneration of a vertebrate limb. It involves cell dedifferentiation and growth. The epidermal cells form a blastema. The blastema gives rise to structures with positional distal values. Retinoic acid changes proximal-distal values in limb regeneration.
Which Organisms can Regenerate?
Regeneration occurs in organisms like hydra, flatworms, tapeworms. They have highly adaptive regenerative capabilities. When an organism is wounded, its cells are activated and the damaged tissues and organs are remodelled back to the original state.
Regeneration is very prominent among metazoans. Starfish, crayfish, reptiles, and amphibians also exhibit signs of tissue regeneration. It is not the same as reproduction. In some animals such as the lizard, the shed limb regrows into the original organ.
Regeneration can happen in many different ways using pluripotent stem cells. Some regeneration does not require stem cells. After amputation, stem cells accumulate at the site of injury. The cells then start dividing to form the missing tissue. But not all animals use the pluripotent cells for regeneration.
Regeneration in Different organisms:
i) Algae
One of the most outstanding feats of regeneration occurs in the single-celled green alga Acetabularia. This plant-like protist of shallow tropical water consists of a group of short rootlike appendages; a long thin “stem,” up to several centimetres in length; and an umbrella-like cap at the top. The entire organism is one cell, with its single nucleus situated at the base in one of the “roots.” If the cap is cut off, a new one regenerates from the healed over stump of the amputated stem. The nucleus is necessary for this kind of regeneration, presumably because it provides the information needed to direct the development of the new cap. Once this information has been produced by the nucleus, however, the nucleus can be removed and regeneration continues unabated.
 |
| Regeneration in lower Plants |
If the nucleus from one species of Acetabularia is added to a cell-body of another species, and the cap of the recipient cell is amputated, the new cap that regenerates will be a hybrid because each nucleus exerts its own morphogenetic influences. On the other hand, if the nucleus from one species is substituted for that in another, regeneration reflects the properties of the new nucleus.
ii) Protozoans
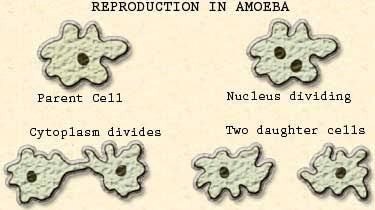
Most single-celled, animal-like protists regenerate very well. If part of the cell fluid, or cytoplasm, is removed from Amoeba, it is readily replaced. A similar process occurs in other protozoans, such as flagellates and ciliates. In each case, however, regeneration occurs only from that fragment of the cell containing the nucleus. Amputated parts that lack a nucleus cannot survive. In some ciliates, such as Blepharisma or Stentor, the nucleus may be elongated or shaped like a string of beads. If either of these organisms is cut in two so that each fragment retains part of the elongated nucleus, each half proceeds to grow back what it lacks, giving rise to a complete organism in less than six hours. The way in which such a bisected protozoan regenerates is almost identical with the way it reproduces by ordinary division. Even a very tiny fragment of the whole organism can regenerate itself, provided it contains some nuclear material to determine what is supposed to be regenerated.
iii) Green plants:
The mechanisms by which vascular plants grow have much in common with regeneration. Their roots and shoots elongate by virtue of the cells in their meristems, the conical growth buds at the tip of each branch. These meristems are capable of indefinite growth, especially in perennial plants. If they are amputated they are not replaced, but other meristems along the stem, normally held in abeyance, begin to sprout into new branches that more than compensate for the loss of the original one. Such a process is called restitution.
Plants are also capable of producing callus tissue wherever they may be injured. This callus is proliferated from cambial cells, which lie beneath the surface of branches and are responsible for their increase in width. When a callus forms, some of its cells may organize into growing points, some of which in turn give rise to roots while others produce stems and leaves.
iv) Coelenterates
The vast majority of research on coelenterates has been focussed on hydras and some of the colonial hydroids. If a hydra is cut in half, the head end reconstitutes a new foot, while the basal portion regenerates a new hydranth with mouth and tentacles. This seemingly straightforward process is deceptively simple. From tiny fragments of the organism whole animals can be reconstituted. Even if a hydra is minced and the pieces scrambled, the fragments grow together and reorganize themselves into a complete whole. The indestructibility of the hydra may well be attributed to the fact that even the intact animal is constantly regenerating itself. Just below the mouth is a growth zone from which cells migrate into the tentacles and to the foot where they eventually die. Hence, the hydra is in a ceaseless state of turnover, with the loss of cells at the foot and at the tips of the tentacles being balanced by the production of new ones in the growth zone. If such an animal is X-rayed, the proliferation of new cells is inhibited and the hydra gradually shrinks and eventually dies owing to the inexorable demise of cells and the inability to replace them.
 |
| Regeneration in Hydra |
In colonial hydroids, such as Tubularia, there is a series of branching stems, each of which bears a hydranth on its end. If these hydranths are amputated they grow back within a few days. In fact, the organism normally sheds its hydranths from time to time and regenerates new ones naturally.
v) Flatworms
Planarian flatworms are well-known for their ability to regenerate heads and tails from cut ends. In the case of head regeneration, some blastema cells become brain tissues, others develop into the eyes, and still others differentiate as muscle or intestine. In a week or so, the new head functions almost as well as the original.
 |
Regeneration in planaria |
The blastema that normally gives rise to a single head is, under certain circumstances, even capable of becoming two heads if the stump of a decapitated flatworm is divided in two by a longitudinal cut. Each of the two halves then gives rise to a complete head. Thus, each blastema develops into an entire structure regardless of its size or position in relation to the rest of the animal.
In the case of flatworms there is still considerable disagreement concerning the origins of the blastema. Some investigators contend that it is derived from neoblasts, undifferentiated reserve cells scattered throughout the body. Others claim that there are no such reserve cells and that the blastema develops from formerly specialized cells near the wound that dedifferentiate to give rise to the blastema cells. Whatever their source, the cells of the blastema are capable of becoming many different things depending upon their location.
Regeneration in flatworms occurs in a stepwise fashion. The first tissue to differentiate is the brain, which induces the development of eyes. Once the head has formed, it in turn stimulates the production of the pharynx. The latter then induces the development of reproductive organs farther back. Thus, each part is necessary for the successful development of those to come after it; conversely, each part inhibits the production of more of itself. If decapitated flatworms are exposed to extracts of heads, the regeneration of their own heads is prevented. Such a complex interplay of stimulators and inhibitors is responsible for the successful regeneration of an integrated morphological structure.
vi)Annelids:
The segmented worms exhibit variable degrees of regeneration. The leeches, as already noted, are wholly lacking in the ability to replace lost segments, whereas the earthworms and various marine annelids (polychaetes) can often regenerate forward and backward. The expression of such regenerative capacities depends very much on the level of amputation. Anteriorly directed regeneration usually occurs best from cuts made through the front end of the worm, with little or no growth taking place from progressively more posterior bisections. Posteriorly directed regeneration is generally more common and extensive. Some species of worms replace the same number of segments as were lost. Hypomeric regeneration, in which fewer segments are produced than were removed, is more common, however.
 |
| Regeneration in Annelids |
Anterior regeneration depends upon the presence of the central nerve cord. If this is cut or deflected from the wound surface, little or no forward regeneration may take place. Posterior regeneration requires the presence of the intestine, removal of which precludes the formation of hind segments. Thus, it would seem that no head will regenerate without a central nervous system, nor a tail without an opening.
vii) Arthropods:
Many insects and crustaceans regenerate legs, claws, or antennas with apparent ease. When insect legs regenerate, the new growth is not visible externally because it develops within the next proximal segment in the stump. Not until the following molt is it released from its confinement to unfold as a fully developed leg only slightly smaller than the original. In the case of crabs, regenerating legs bulge outward from the amputation stump. They are curled up within a cuticular sheath, not to be extended until the sheath is molted. Lobsters and crayfish regenerate claws and legs in a straightforward manner as direct outgrowths from the stumps. As in other crustaceans, however, these regenerates lie immobile within an enveloping cuticle and do not become functional until their sheath is shed at the next molt.
 |
| Regeneration in insects |
In all arthropods regeneration is associated with molting, and therefore takes place only during larval or young stages. Most insects do not initiate leg regeneration unless there remains ample time prior to the next scheduled molt for the new leg to complete its development. If amputation is performed too late in the intermolt period, the onset of regeneration is delayed until after shedding; the regenerate then does not appear until the second molt. Metamorphosis into the adult stage marks the end of molting in insects, and adults accordingly do not regenerate amputated appendages.
ix) Crustaceans :
Crustaceans often tend to molt and grow throughout life. They therefore never lose the ability to grow back missing appendages. When a leg is lost, a new outgrowth appears even if the animal is not destined to molt for many months. Following a period of basal growth, during which a diminutive limb is produced, the regenerated part eventually ceases to elongate. Not until a few weeks before the next molt does it resume growth and complete its development, triggered by the hormones that induce molting.
x)Fishes:
Many different parts of the fish’s body will grow back. Plucked scales are promptly replaced by new ones, and amputated gill filaments can regenerate easily. The “whiskers,” or taste barbels, of the catfish grow back as perfect replicas of the originals. The most conspicuous regenerating structures in fishes, however, are the fins. When any of these are amputated, new fins grow out from the stumps and soon restore everything that was missing. Even the coloured stripes or spots that adorn some fins are reconstituted by new pigment cells that repopulate the regenerated part. Fin regeneration depends on an adequate nerve supply. If the nerves are cut leading into the fin, regeneration of neither the amputated fin nor excised pieces of the bony fin rays can take place.
 |
| regeneration of fins |
xii) Amphibians:
Salamanders are remarkable for their ability to regenerate limbs. Larval frogs, or tadpoles, also possess this ability, but usually lose it when they become frogs. It is not known why frog legs do not regenerate, and under appropriate stimuli they can be induced to do so.
Tadpoles and salamanders can replace amputated tails. Tadpole tails have a stiff rod called the notochord for support, whereas salamanders possess a backbone, composed of vertebrae. Both tails contain a spinal cord. When the salamander regenerates its tail, the spinal cord grows back and segmental nerve-cell clusters (ganglia) differentiate. Tadpoles also regenerate their spinal cords, but not the associated ganglia. If the spinal cord is removed or destroyed in the salamander, no tail regeneration occurs; if it is removed from the tadpole tail, however, regeneration can proceed without it.
xii) Reptiles:
Lizards also regenerate their tails, especially in those species that have evolved a mechanism for breaking off the original tail when it is grasped by an enemy. When the lizard tail regenerates, however, it does not replace the segmented vertebrae. Instead, there develops a long tapering cartilaginous tube within which the spinal cord is located and outside of which are segmented muscles. The spinal cord of the lizard tail is necessary for regeneration, but the regenerated tail does not reproduce the ganglia that are normally associated with it. Occasionally, a side tail may be produced if the original tail is broken but not lost.
xiii) Birds:
Regeneration of amputated appendages in birds is not known to occur; however, they do replace their feathers as a matter of course. While most species shed and regenerate feathers one at a time so as not to be grounded, flightless birds, such as penguins, may molt them all at once. Male puffins cast off their colorful beaks after the mating season, but grow new ones the following year. In like manner, the dorsal keel on the upper beaks of male pelicans is shed and replaced annually.
 |
| Regeneration of birds |
ix) Mammals:
Although mammals are incapable of regenerating limbs and tails, there are a few exceptional cases in which lost tissues are in fact regenerated. Not the least of these cases is the annual replacement of antlers in deer. These remarkable structures, which normally grow on the heads of male deer, consist of an inner core of bone enveloped by a layer of skin and nourished by a copious blood supply. During the growing season the antlers elongate by the proliferation of tissues at their growing tips. The rate of growth in some of the larger species may surpass one centimetre (0.39 inch) per day; the maximum rate of growth recorded for the elk is 2.75 centimetres (1.05 inches) per day. When the antlers have reached their full extent, the blood supply is constricted, and the skin, or velvet, peels off, thus revealing the hard, dead, bony antlers produced by the male deer in time for the autumn mating season. The regeneration of elk antlers spans about seven months. The following spring, the old antlers are shed and new ones grow to replace them.
Still another example of mammalian regeneration occurs in the case of the rabbit’s ear. When a hole is punched through the external ear of the rabbit, tissue grows in from around the edges until the original opening is reduced or obliterated altogether. This regeneration is achieved by the production of new skin and cartilage from the margins of the original hole. A similar phenomenon occurs in the case of the bat’s wing membrane.
Regeneration in Humans:
The regrowth of lost tissues or organs in the human body is being researched. Some tissues such as skin regrow quite readily; others have been thought to have little or no capacity for regeneration, but ongoing research suggests that there is some hope for a variety of tissues and organs .Human organs that have been regenerated include the bladder, vagina and the penis.
As are all metazoans, humans are capable of physiological regeneration (i.e. the replacement of cells during homeostatic maintenance that does not necessitate injury). For example, the regeneration of red blood cells via erythropoiesis occurs through the maturation of erythrocytes from hematopoietic stem cells in the bone marrow, their subsequent circulation for around 90 days in the blood stream, and their eventual cell-death in the spleen. Another example of physiological regeneration is the sloughing and rebuilding of a functional endometrium during each menstrual cycle in females in response to varying levels of circulating estrogen and progesterone.

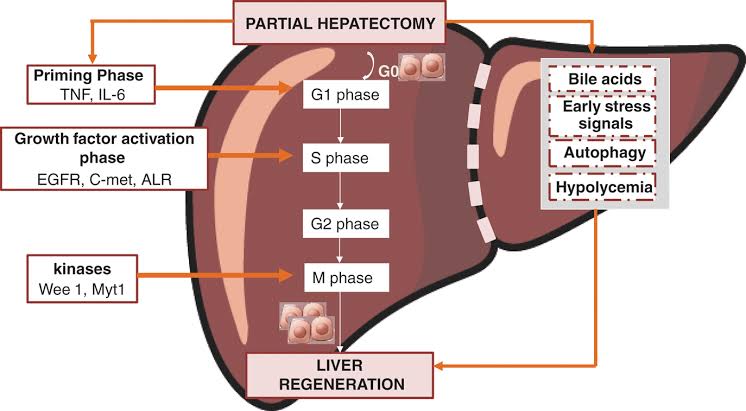
However, humans are limited in their capacity for reparative regeneration, which occurs in response to injury. One of the most studied regenerative responses in humans is the hypertrophy of the liver following liver injury. For example, the original mass of the liver is re-established in direct proportion to the amount of liver removed following partial hepatectomy,which indicates that signals from the body regulate liver mass precisely, both positively and negatively, until the desired mass is reached. This response is considered cellular regeneration (a form of compensatory hypertrophy) where the function and mass of the liver is regenerated through the proliferation of existing mature hepatic cells (mainly hepatocytes), but the exact morphology of the liver is not regained. This process is driven by growth factor and cytokine regulated pathways.
 |
Regeneration in liver |
Adult neurogenesis is also a form of cellular regeneration. Even in adult myocardium following infarction, proliferation is only found in around 1% of myocytes around the area of injury, which is not enough to restore function of cardiac muscle. However, this may be an important target for regenerative medicine as it implies that regeneration of cardiomyocytes, and consequently of myocardium, can be induced.
Another example of reparative regeneration in humans is fingertip regeneration
Yet another example of regeneration in humans is vas deferens regeneration, which occurs after a vasectomy and which results in vasectomy failure.
(note: I tried to present all important points, facts, examples of some of the phyllum in the animal kingdom very briefly and with a good accuracy)~ the author, vetuniverse.blogspot.com
This article is Written / Authored by : Sahidul Islam on 1st dec, 2020
(All copyrights belongs to vetuniverse.blogsopt.com)
(No part of this article should be modified, reproduced, changed, reformed, copied in any forms without the proper permission from the author )




👍👍👍
ReplyDeleteThank you For letting me know that This article found your interest. Keep reading.🤗👍🏼
Delete